 中文网站
中文网站
Током ПЦР реакције, често се сусрећу неки фактори који ометају.
Због веома високе осетљивости ПЦР-а, контаминација се сматра једним од најважнијих фактора који утичу на резултате ПЦР-а и може произвести лажно позитивне резултате.
Подједнако критични су и различити извори који доводе до лажно негативних резултата. Ако су један или више битних делова PCR смеше или саме реакције амплификације инхибирани или ометани, дијагностички тест може бити отежан. То може довести до смањене ефикасности, па чак и лажно негативних резултата.
Поред инхибиције, губитак интегритета циљне нуклеинске киселине може се десити због услова транспорта и/или складиштења пре припреме узорка. Посебно, високе температуре или неадекватно складиштење могу довести до оштећења ћелија и нуклеинских киселина. Фиксација ћелија и ткива и уграђивање у парафин су добро познати узроци фрагментације ДНК и стални проблем (видети слике 1 и 2). У овим случајевима, чак ни оптимална изолација и пречишћавање неће помоћи.
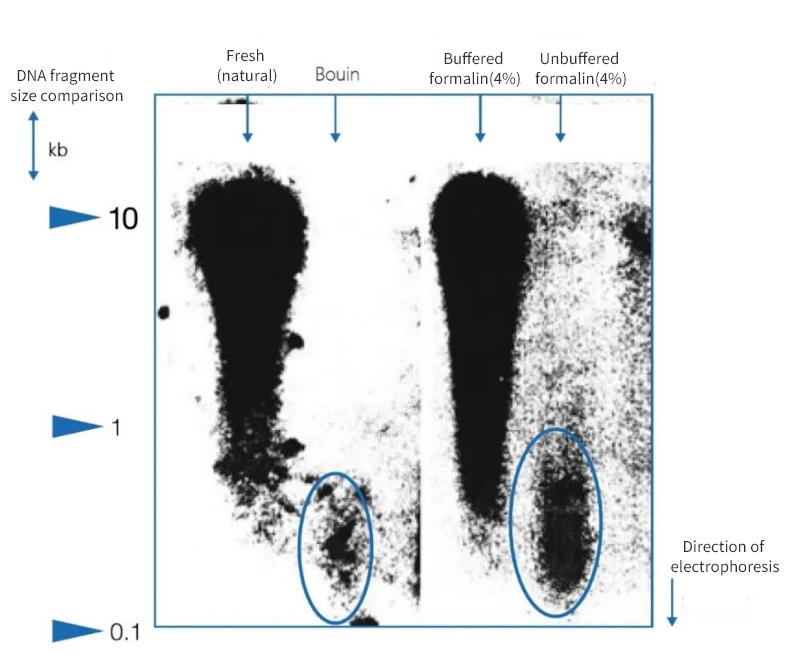
Слика 1 | Утицај имобилизације на интегритет ДНК
Електрофореза на агарозном гелу показала је да се квалитет ДНК изоловане из парафинских пресека аутопсија значајно разликује. ДНК различитих просечних дужина фрагмената била је присутна у екстрактима у зависности од методе фиксације. ДНК је сачувана само када је фиксирана у нативним замрзнутим узорцима и у пуферованом неутралном формалину. Употреба јако киселог Буеновог фиксатива или непуферованог формалина који садржи мрављу киселину довела је до значајног губитка ДНК. Преостала фракција је високо фрагментирана.
Са леве стране, дужина фрагмената је изражена у килобазним паровима (kbp)
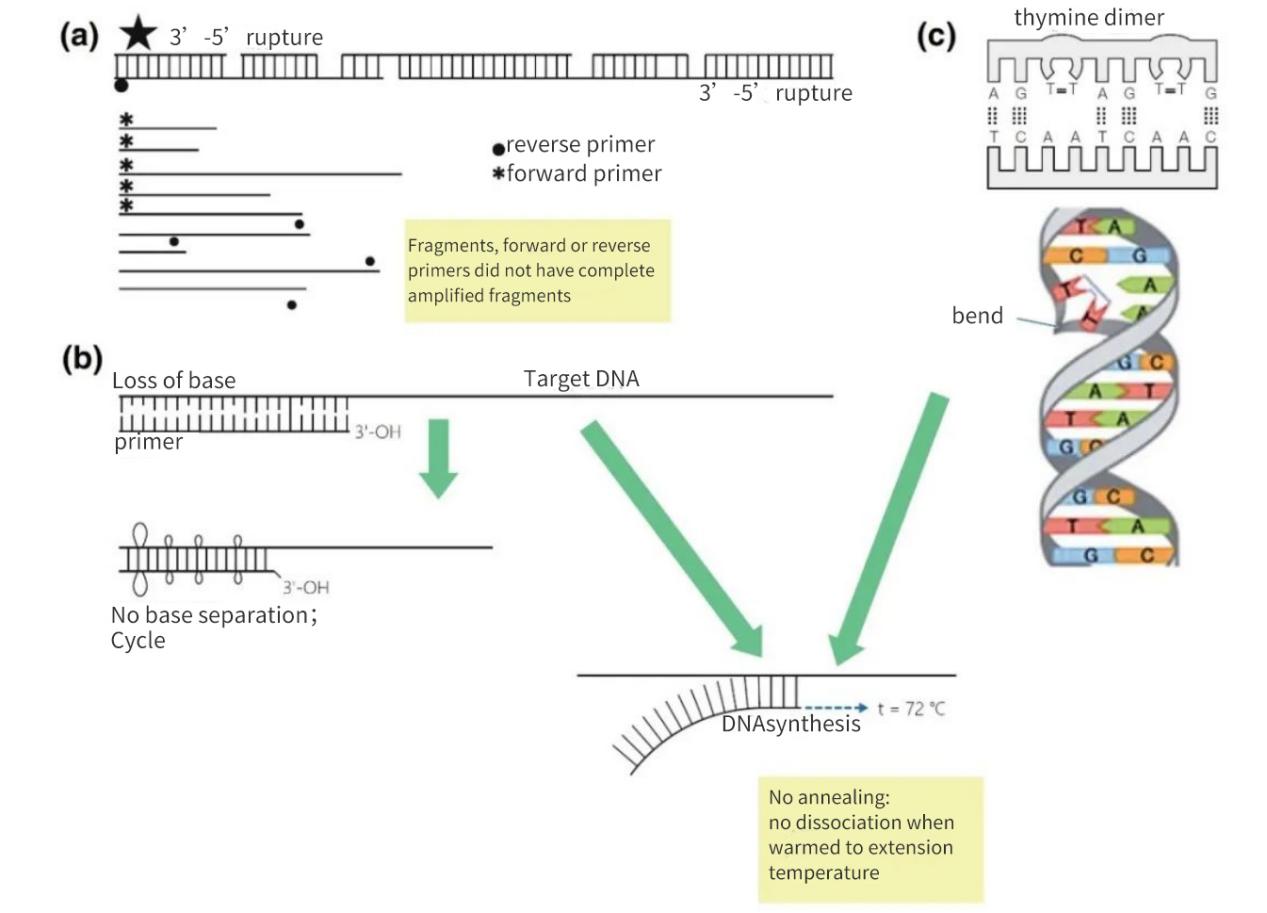
Слика 2 | Губитак интегритета циљева нуклеинских киселина
(а) Размак од 3′-5′ на оба ланца резултираће прекидом у циљној ДНК. Синтеза ДНК ће се и даље одвијати на малом фрагменту. Међутим, ако на фрагменту ДНК недостаје место за везивање прајмера, долази само до линеарне амплификације. У најповољнијем случају, фрагменти се могу поново заситити, али ће приноси бити мали и испод нивоа детекције.
(б) Губитак база, углавном због депуринације и формирања тимидинског димера, доводи до смањења броја водоничних веза и смањења температуре загревања (Tm). Током продужене фазе загревања, прајмери ће се истопити од матричне ДНК и неће се жарити чак ни под мање строгим условима.
(ц) Суседне тиминске базе формирају ТТ димер.
Још један уобичајени проблем који се често јавља у молекуларној дијагностици јесте мање него оптимално ослобађање циљних нуклеинских киселина у поређењу са екстракцијом фенол-хлороформом. У екстремним случајевима, ово може бити повезано са лажно негативним резултатима. Много времена се може уштедети лизом кључањем или ензимском дигестијом ћелијских остатака, али ова метода често резултира ниском осетљивошћу ПЦР због недовољног ослобађања нуклеинских киселина.
Инхибиција полимеразне активности током амплификације
Генерално, инхибиција се користи као општи концепт за описивање свих фактора који доводе до субоптималних резултата PCR. У строго биохемијском смислу, инхибиција је ограничена на активност ензима, тј. смањује или спречава конверзију супстрата у производ кроз интеракцију са активним местом ДНК полимеразе или њеним кофактором (нпр. Mg2+ за Taq ДНК полимеразу).
Компоненте у узорку или различити пуфери и екстракти који садрже реагенсе могу директно инхибирати ензим или везати његове кофакторе (нпр. EDTA), чиме инактивирају полимеразу и заузврат доводе до смањених или лажно негативних PCR резултата.
Међутим, многе интеракције између компоненти реакције и нуклеинских киселина које садрже мету се такође називају „инхибитори ПЦР-а“. Када се интегритет ћелије наруши изолацијом и нуклеинска киселина се ослободи, могу доћи до интеракција између узорка и његовог околног раствора и чврсте фазе. На пример, „хватачи“ могу да се вежу за једноланчану или дволанчану ДНК путем нековалентних интеракција и да ометају изолацију и пречишћавање смањењем броја мета које на крају стигну до реакционе посуде за ПЦР.
Генерално, инхибитори ПЦР-а присутни су у већини телесних течности и реагенса који се користе за клиничке дијагностичке тестове (уреа у урину, хемоглобин и хепарин у крви), дијететским суплементима (органске компоненте, гликоген, масти, Ca2+ јони) и компонентама у животној средини (феноли, тешки метали).
| Инхибитори | Извор |
| калцијум јони | Млеко, коштано ткиво |
| Колаген | Тканино |
| Жучне соли | Фецес |
| Хемоглобин | У крви |
| Хемоглобин | Узорци крви |
| Хуминска киселина | Земљиште, биљка |
| Крв | Крв |
| Лактоферин | Крв |
| (Европски) меланин | Кожа, коса |
| Миоглобин | Мишићно ткиво |
| Полисахариди | Биљка, измет |
| Протеаза | Млеко |
| Уреа | Урин |
| Мукополисахарид | Хрскавица, слузокоже |
| Лигнин, целулоза | Биљке |
Више распрострањених инхибитора ПЦР-а могу се наћи у бактеријама и еукариотским ћелијама, ДНК која није циљна, макромолекулима ткивних матрица које се везују за ДНК и лабораторијској опреми као што су рукавице и пластика. Пречишћавање нуклеинских киселина током или након екстракције је пожељна метода за уклањање инхибитора ПЦР-а.
Данас, различита аутоматизована опрема за екстракцију може заменити многе ручне протоколе, али 100% опоравак и/или пречишћавање циљева никада није постигнуто. Потенцијални инхибитори могу и даље бити присутни у пречишћеним нуклеинским киселинама или су можда већ ступили на снагу. Постоје различите стратегије за смањење утицаја инхибитора. Избор одговарајуће полимеразе може имати значајан утицај на активност инхибитора. Друге доказане методе за смањење инхибиције ПЦР су повећање концентрације полимеразе или примена адитива као што је БСА.
Инхибиција PCR реакција може се показати употребом интерне контроле квалитета процеса (IPC).
Мора се водити рачуна да се сви реагенси и други раствори у комплету за екстракцију, као што су етанол, EDTA, CETAB, LiCl, GuSCN, SDS, изопропанол и фенол, уклоне из изолата нуклеинске киселине темељним прањем. У зависности од њихове концентрације, они могу активирати или инхибирати PCR.
Време објаве: 19. мај 2023.
 Подешавања приватности
Подешавања приватности